
Biophysical Research on Biomaterials for Biomimetics
NASA URETI BiMAT Projects
Bone | Diatoms | Mussel and Nereis Worm | Spider silk | Tendon collagen-1
Introduction
In 2002, a new institute was formed, of which four collaborating groups at UCSB - the Paul Hansma, Dan Morse [Marine Science; Molecular, Cellular and Developmental Biology], the Galen Stucky Group [Chemistry] and the Herb Waite Group [Marine Science] - were a part. This institute, the NASA University Resarch, Engineering and Technology Institute on Biologically Inspired Materials (BiMAT), was formed as a five-year initiative to develop advanced biomimetic materials for NASA's aerospace vehicles.
Our group has researched and published papers on an array of biomaterials in this collaboration so far, particularly with the Morse, Stucky and Waite groups - bone, spider-web capture-silk threads, diatom adhesives, collagen-1 in tendon, mussel byssus and the jaws of the Nereis worm. The common denominator that interests us in all these materials is their amazing strength given their light weight. The complexity and sophistication of their hierarchical structures is what makes these biomaterials extremely tough. Because they are living, they can also heal themselves in a dynamic fashion. The goal of the BiMAT Institute is to convert fundamental discoveries that its reserach group members produce into advanced, tough, and self-healing space materials.
Bone
Please visit our page on our bone research.
Diatom glue
Diatoms are unicellular microalgae with a cell wall consisting of a siliceous skeleton enveloped by an organic case essentially composed of polysaccharides and proteins. The cell walls form a pillbox-like shell (called the frustule) consisting of two valves that fit within each other with the help of a set of girdle bands. Frustules vary greatly in shape, ranging from box-shaped to cylindrical; they can be symmetrical as well as asymmetrical and exhibit an amazing diversity of nanostructured frameworks (Figure 1).
Diatoms are found in both freshwater and marine environments, as well as in moist soils, and on moist surfaces. They are either freely floating (planktonic forms) or attached to a substrate (benthic forms), and some species may form chains of cells of varying length. Individual diatoms range from 2µm up to several millimetres in size, although only few species are larger than 200µm. Diatoms as a group are very diverse with 12,000– 60,000 species reported.
Currently, human chemical synthesis cannot produce siliceous structures with the hierarchical structural detail of the diatom frustules nor can ordered siliceous structures be produced synthetically under the benign conditions of diatom biomineralization. Biosilicification occurs at ambient temperatures and pressures, whereas artificial chemical synthesis of silica-based materials (e.g. resins, molecular sieves and catalysts) requires extreme conditions of temperature, pressure and pH.

Figure 1 Diatoms are unicellular organisms whose surface comprises nanostructured amorphous glass. Scanning electron micrographs of frustules of Ellerbeckia arenaria, a freshwater diatom, reveal amazingly beautiful and diverse details on a single cell.
Engineering stable underwater adhesives currently poses a major technical challenge, because most man-made adhesives fail in wet conditions, owing to chemical modification of the adhesive or its substrate. But diatoms produce adhesives that are extremely strong and robust, in both fresh- and seawater environments. In our AFM study of living diatoms, our phase imaging and force spectroscopy experiments revealed characteristics of these natural adhesives that may be of use in designing man-made adhesive analogues that function in wet environments. [Gebeshuber, I. C. et al. Atomic force microscopy study of living diatoms in ambient conditions. Journal of Microscopy-Oxford 212 , 292-299 (2003)].
The AFM allows for investigations of micromechanical properties of the cell surface, for example viscoeleastic properties, adhesion forces and hardness measurements, in physiological conditions. Effortless sample preparation and unprecedented resolution on insulating materials are other benefits. In our study on living diatoms, we experimented with three diatom species that produce outstanding natural adhesives: Eunotia sudetica, Navicula seminulum and an unidentified species.
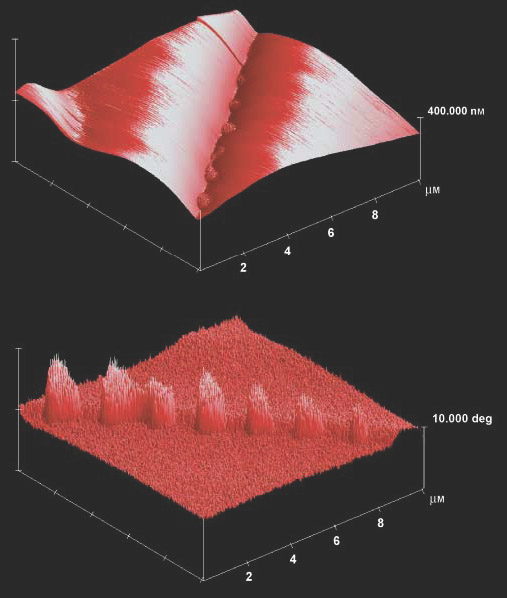
Figure 2 AFM topography and phase images of the glue that sticks cells of the diatom Eunotia sudetica together. Top: Topography image of glue clumps between cells with undulating surfaces. Bottom: AFM phase images convert the phase delay between the drive and response of the cantilever into an image. Phase images contain information about the energy dissipated during the interaction of the AFM tip with the sample, and can help us to understand the viscoelastic and adhesion properties of the surfaces investigated - here, the properties of the organic material responsible for diatom adhesion. The diatom adhesive causes a phase lag of about 10° compared with the rest of the frustule surfaces, where on a single frustule it is within 1°. Note the 2° interfrustule phase step, which reveals slightly different viscoelastic properties of the two neighbouring valves. Note that for better view, (b) is rotated clockwise by 90 ° compared with (a).
Toughness mechanisms in marine organisms: mechanical gradients
Tremendous mechanical stresses can be experienced in the area at the interface of the hard and soft tissues that comprise most organisms. Little is known about how nature works to mitigate these stresses; our studies on two noncellular tissues, mussel byssus and polychaete jaws suggest that one natural strategy to relieve interfacial stresses between adjoining stiff and soft tissue appears to be the creation of a fuzzy boundary, which avoids abrupt changes in mechanical properties, and graduates the transcendence from stiff to soft.
In manufacturing synthetic materials, two methods can be employed to mitigate interfacial stresses: (1) increasing the energy of interaction across the contact zone between areas of different mechanical properties; or (2) avoiding sharp boundaries between materials. Although the first option may increase the stress at failure, it does not decrease interfacial stresses. It has been practiced for many years and is the basis for priming or surface coupling treatments of glass or metals that precede bonding with polymers. The second option, which has surfaced only recently, is the manufacture of functional gradients. In other words, if material A can be processed so that it gradually or incrementally becomes material B (and vice versa), then interfacial stresses can be dissipated over a much larger surface area and volume.
The byssus is a connective tissue peculiar to mussels, and is remarkable because, although it is remarkably stretchy, it is also stiff and strong. It is deposited outside the confines of the soft living mussel tissue and contains no cells for maintenance or repair. Byssus securely attaches mussels to rocks and pilings against the strong, repetitive forces generated by waves. The byssus thus mediates between a very stiff inert material (e.g., rock) and very soft living tissue (e.g. the mussel). Because mussel byssus is a bundle of several hundred threads, each measuring between 2 and 4 cm in length and 50 µ m in diameter (in Mytilus edulis and M. gallopro vincialis), every individual thread represents a unit of attachment with the distal end bonded to rock and proximal end inserted into living mussel tissue. Previous scanning electron microscopy and biomechanical studies have established that it is much more than a tether but represents a mechanically graded fiber that is significantly stiffer at the distal end where it joins to rocks than at the proximal end where it joins to living tissue.
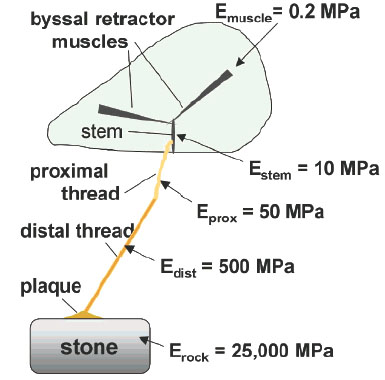
Figure 3 Schematic mussel on the half-shell with one byssal thread showing the incremental steps in stiffness, E, from the retractor muscles to the rock. Note the 10-fold decrease in stiffness between the distal and proximal portions of the thread. (Adapted from Kornberg, A. (2003) Sci. 28, 515-517.).
Much recent work has focused on the fibrous proteins that make up the different portions of the byssal thread. The simplest unit of structure is a trimer of preCOL chains (~80 kD each). We characterized the proteins composing byssal threads, their locations along the thread, their mechanical and chemical properties, and discussed their contributions to the graduating mechanical properties of the byssal thread [Waite, J. H., Lichtenegger, H. C., Stucky, G. D. & Hansma, P. Exploring molecular and mechanical gradients in structural bioscaffolds. Biochemistry 43 , 7653-7662 (2004)].
Herb Waite's previous work on the collagen containing threads from both M. edulis and M. gallopro v incialis showed that ultimate or breaking strain is between 100 and 200% in the proximal portion and is an important feature of thread toughness [Waite, J. H. et al. Matrix Biology 17, 93-106 (1998). This strain greatly exceeds the ultimate strain of ~10% for more typical collagenous structures such as tendon [Gosline, J. et al. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 357, 121-132 (2002) - (this is not a Hansma Group paper)]. Toughness in a variety of biological materials seems to be linked to the reversible sacrificial unfolding of modular domains to provide extra length during deformation, for example in titin.
We dissected individual byssal threads from intact byssus shed by marine mussels (Mytilus gallopro vincialis), and imaged them with an Atomic Force Microscope (Digital Instruments Nanoscope IIIa, Digital Instruments, Santa Barbara, CA), on which images were recorded at a scan rate of 1 Hz using standard silicon tapping-mode tips (force constant 40 N/m, resonant frequency 300 kHz), at room temperature in air. Our examination of preCOL-NG assemblies in the proximal thread revealed many levels of hidden extra length that may contribute to the extraordinary observed strain.
Although much work has been done on the structure and formation of byssal threads, the specific shape of mesogenic units, the nature of their ordered assembly in the byssus, and how they deform under tension are still unknowns. We worked on characterizing the gigantic banana-shaped protein mesogens used by marine mussels to make their byssal threads, and determined that the sacrificial bonds and hidden length system in these protein molecules contributes to the amazing strains the threads can withstand before yielding [Hassenkam, T., Gutsmann, T., Hansma, P., Sagert, J. & Waite, J. H. Giant bent-core mesogens in the thread forming process of marine mussels. Biomacromolecules 5 , 1351-1355 (2004)].
It is quite likely that nature has many other examples of fibers and films based on giant bent-core mesogens. Fundamental studies of these could provide exciting bio-inspired design paradigms for a host of new materials properties and applications.
We also studied the well-developed jaws of the Nereis marine worm, which uses the jaw to grasp, inject venom into, and dismantle prey. The jaws contain proteins with histidine-rich domains and transition metals, Cu and Zn, and, in nanoindentation experiments, we confirmed the distinct mechanical gradients the jaws display, with the jaw tips being much stiffer than the jaw base. We investigated the chemical make-up of the jaws, particularly the basis for the preference mussels and worms have for using histidine-metal groups to stabilize the structural proteins that make up most of the jaw, instead of covalently cross-linking them. Histidine-metal interactions can be robustly and instantly formed, for example by being triggered to be formed at pH changes; they are also reversible and so can self-heal. The addition of only a few percent Zn ions with the jaw proteins makes it 2-3 times harder and stiffer than the best organic synthetic polymers, and ranks it near dentin in hardness, even though dentin is over 70% mineral by weight.
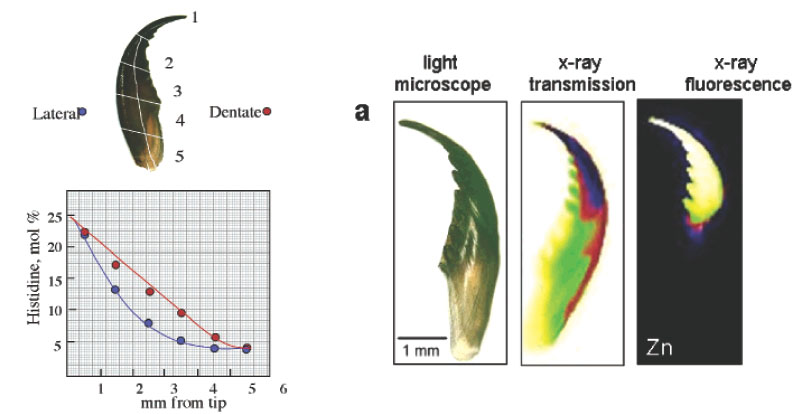
Figure 4 Gradients in the tip-to-base histidine and zinc distribution of a Nereis jaw (from Pender et al. Acta Mater. 49, 3255-3262 - not a Hansma Group paper). The left panel shows a sketch of a Nereis jaw sectioned for amino acid analysis (top) and the decrease of histidine content (bottom) from ~22 mol % at the tip to 5 mol % at the base and lateral to serrated side of the Nereis jaw. The right panel shows the Zn-fluorescence mapping with synchrotron radiation. Images from left to right display a light microscopic image of intact Nereis jaw, an X-ray absorption image (the dark region at the tip denotes greater absorption), and an X-ray fluorescence image (lighter colors indicate greater amount of Zn).
Our papers in this area so far are:
Hassenkam, T., Gutsmann, T., Hansma, P., Sagert, J. & Waite, J. H. Giant bent-core mesogens in the thread forming process of marine mussels. Biomacromolecules 5 , 1351-1355 (2004).
Waite, J. H., Lichtenegger, H. C., Stucky, G. D. & Hansma, P. Exploring molecular and mechanical gradients in structural bioscaffolds. Biochemistry 43 , 7653-7662 (2004).
Spiderweb capture-silk threads
In order to learn why spider web silk threads are so remarkably strong and elastic, we studied spider web capture-silk threads: the very sticky, strong and elastic spiraled material in the webs of orb-weaving spiders. We studied them with AFM force spectroscopy experiments, as force spectroscopy enables the mechanical testing of capture silk alone, which is otherwise difficult to separate from dragline silk. [Becker, N. et al. Molecular nanosprings in spider capture-silk threads. Nature Materials 2, 278-283 (2003)].
After force spectroscopy experiments with the Atomic Force Microscope, pulling bulk threads, and amino acid sequencing, we we able to present initial models for capture silk's molecular and multimolecular structure. We had found that spider capture silk requires exponential force increases as it is pulled (In each capture-silk pull, the successive rupture peaks usually occurred at increasing forces). Intact spider dragline silk does not show an exponential force increase when stretched. We also found evidence that capture silk molecules contain sacrificial bonds and hidden length that reform quickly if the protein is allowed to relax after it is extended. We proposed two models for why the pulling force increased exponentially: simply, as intact capture silk is composed of an assembly of molecules, either that the molecules successively rupture or detach, sometimes even in multiple steps because of looping, or that the molecular assembly is networked by crosslinking springs. We discussed the possible nature of the cross-linking springs, proposing that they may be a part of the glue that is known to coat capture-silk.
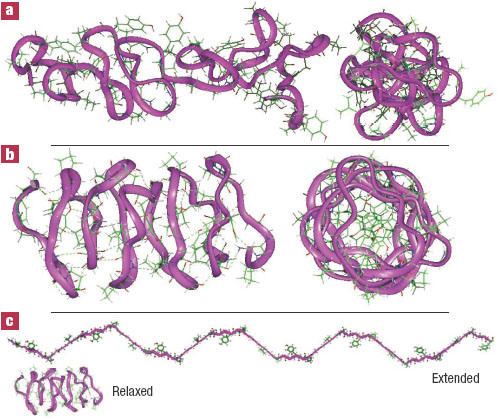
Figure 5 Our β-spiral models for relaxed and extended protein conformations in spider capture silk - side and end views. a) Araneus gemmoides, 85 amino acids long, in its relaxed state. b) Nephila clavipes, 75 amino acids long - relaxed state. c) Nephila clavipes stretched to its maximum extension without deforming bond angles. The capture silk from both these species of spider contain high percentages of glycine and proline, often in the repetitive GPGGXn motifs characteristic of β-spirals.
Please click here [LINK "HERE" to PDF] for the paper reporting this project.
Collagen in tendon
We have also investigated structure-function relationships and mechanical properties of collagen, primarily the collagen of rat-tail tendon fibrils. We studied tendons because they are among the highest stress-carrying structures in mammals. Compared with other structural proteins, such as spectrin, tenascin and spider silk protein, the forces needed to stretch tendon molecules are high. Tendon is made up of ~90% collagen, 10% proteoglycans and glycoproteins, and are primarily elastic.
We have published three papers in this area so far:
Venturoni, M., Gutsmann, T., Fantner, G. E., Kindt, J. H. & Hansma, P. K. Investigations into the polymorphism of rat tail tendon fibrils using atomic force microscopy. Biochemical and Biophysical Research Communications 303, 508-513 (2003).
Gutsmann, T. et al. Evidence that collagen fibrils in tendons are inhomogeneously structured in a tubelike manner. Biophysical Journal 84, 2593-2598 (2003).
Gutsmann, T. et al. Force spectroscopy of collagen fibers to investigate their mechanical properties and structural organization. Biophysical Journal 86, 3186-3193 (2004).
In the exploratory study Venturoni et al., we discovered, apart from the well-known 67nm banding periodicity in type 1 collagen, two banding patterns with 23 ± 2 nm and 210 ± 15nm periodicities. We introduced a discussion on these periodicities for further investigation in light of synergistic research. Interestingly, we also observed that although collagen fibrils dissolve in a pH of less than 4, the presence of salt ions (Na+ and Ca+ here) prevented this dissolution.

Figure 6 Deflection image of a bundle of dried collagen fibrils, showing (a) a collagen fibril having a periodicity of about 23nm and (b) a fibril with a typical spacing of about 67nm.
In the first Gutsmann et al paper, we reported that we observed with AFM that collagen fibrils act as inhomogenously packed and structured tubes rather than closely-packed, homogenously structured rods. This may be significant because common models of tendon assume collagen fibrils are laterally and homogenously packed. We also observed that collagen fibrils are not hollow, but possess a structure with the shell and core having different mechanical properties. The shell, we saw, is harder than the core. The fibrils can fold back onto themselves, make very tight turns or ‘‘kinks,’’ become flat as they are squeezed, and wrap around one another. They shrink when they are dried, because their softer core contracts much more than their harder shell. Our data suggested that collagen molecules are more highly crosslinked near the fibril surface and more disordered or soft in the central region. Our observations are consistent with x-ray diffraction data, which has shown collagen molecules in rat-tail tendons appearing to be in a state of liquidlike disorder (Fratzl et al., 1993; Hulmes et al., 1995; Prockop and Fertala, 1998), nuclear magnetic resonance data indicating that the collagen backbone is free to reorient within a fibril (Trochia, 1982; Sarkar et al., 1985), and other prior observations of collagen structure.

Figure 7 AFM images (amplitude signal) of dried collagen fibrils. These images show that banding continues right up to abrupt ends, suggesting that a banded, relatively rigid tube is fractured to expose a more flexible core material.
The importance of our force spectroscopy project [Biophysical Journal 86, 3186-3193 (2004)] was our correlation of the periodic patterns in the topography of rat tail tendon collagen fibrils with the binding patterns observed in our force spectroscopy experiments between collagen fibril molecules. We used small cantilevers to pull subunits out of collagen fibrils of native rat tail tendon. In this study, we probably only studied the mechanical properties of the outer shell.
We found that collagen molecules can rupture sacrificial bonds at constant values of the distances they are stretched. We observed two patterns of rupture: After applying forces greater than 500pN, several ruptures would occur for every 78 ± 3 nm the molecule was stretched. (It takes forces of over 1000pN to break a collagen backbone). This never occurred more than 5 times in the hundreds of molecules stretched. This means that there are bonds between molecules in the fibrils 78nm apart. The other pattern of weaker bonds was barely detectable because the jumps in force applied were only up to 7pN. The ruptures occurred at the periodicity of 22 ± 2 nm, and occurred in the linear-like region of the force-distance curve.
We think the first pattern occurs when bundles of collagen molecules are pulled together, and individual subunits then rupture sequentially. The peak force at each successive rupture decreased probably because after the backbone of one collagen molecule yields, less force is needed to rupture the remaining molecules. The 78nm periodicity is ~16% larger than the characteristic 67nm banding periodicity in wet collagen. It occurred here because the wavy collagen was projected and stretched over an x-y plane, and because the collagen molecules are tilted against the fibril's axis. A direct correlation between the topography of the collagen fibrils and their force spectra. Based on our data, we proposed in our paper that one distinct distance between two binding or cross-linking positions between stretched tropocollagen units is 78nm.
The small jumps in force of < 7pN in the linear-like increase region of the force-extension curves are remarkable because they modulate the increase in force. They also correlate to a smaller banding pattern we observed at a periodicity of 23nm along the collagen fibrils, and are probably due to a gliding effect within the fibrils. These cross-linked regions are probably caused by electrostatic interactions between the molecules in the fibrils, and other forces. The presence of the hidden length mechanism, with its three-dimensional structure, is unlikely.
Typically, these bonds did not reform, unlike other modular proteins with compact domains, such as titin.
We propose both types of the bonds we observed occur in all collagen fibrils, as significantly different forces are needed to break them. Built-in molecular bonds of different strengths and periodicities could contribute to the optimization of the mechanical properties of new materials.